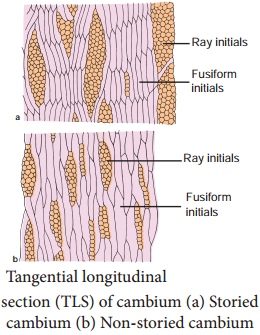
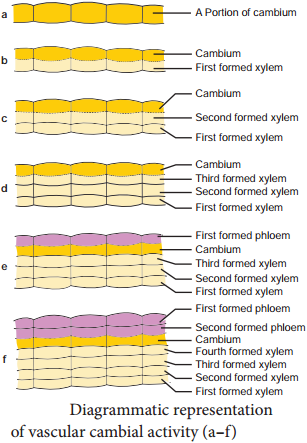
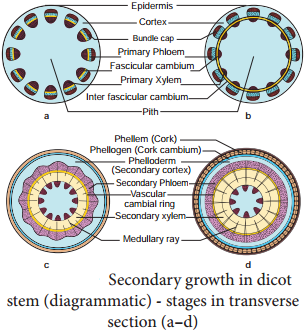
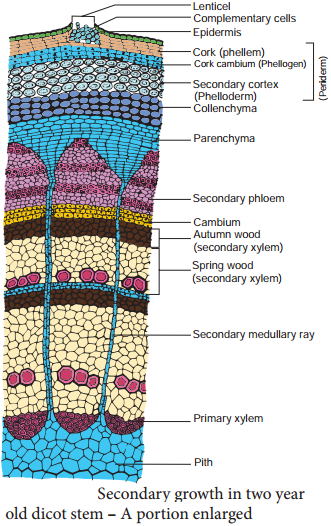
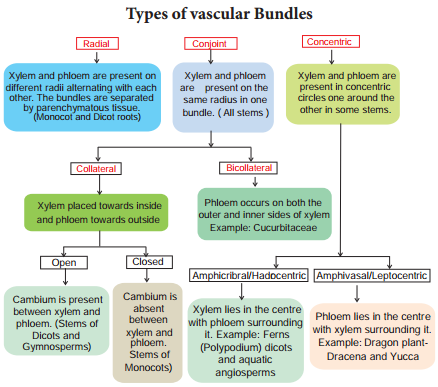
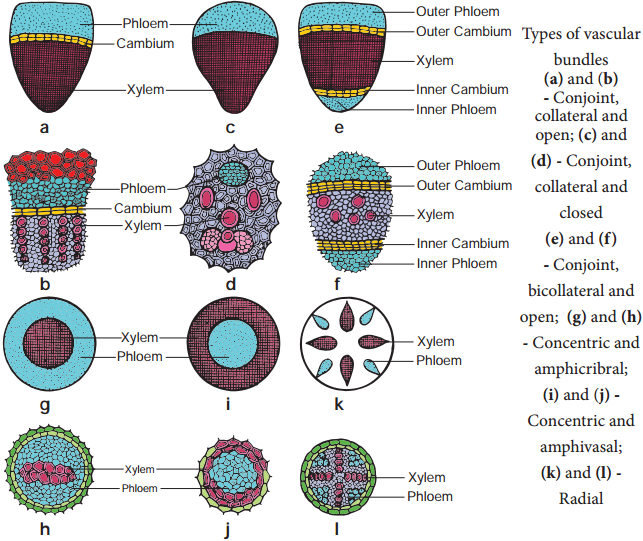
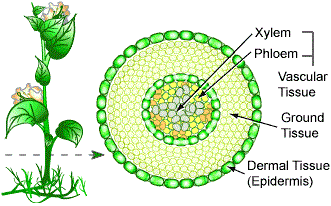
Learninsta presents the core concepts of Biology with high-quality research papers and topical review articles.
Secondary Growth in Dicot Root
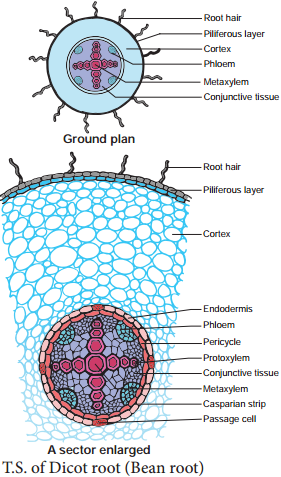
Secondary growth in dicot roots is essential to provide strength to the growing aerial parts of the plants. It is similar to that of the secondary growth in dicot stem. However, there is marked diffrence in the manner of the formation of vascular cambium.
The vascular cambium is completely secondary in origin. It originates from a combination of conjunctive tissue located just below the phloem bundles, and as a portion of pericycle tissue present above the protoxylem to form a complete and continuous wavy ring. This wavy ring later becomes circular and produces secondary xylem and secondary phloem similar to the secondary growth in stems.
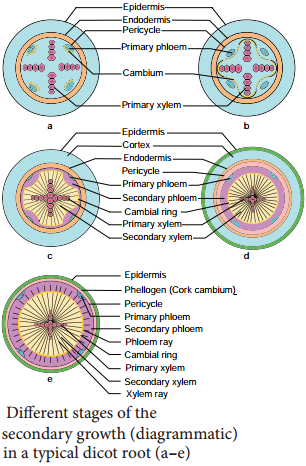
Differences Between Secondary Growth in Dicot Stem and Root
Secondary Growth in Dicot Stem | Secondary Growth in Dicot Root |
| 1. The cambial ring formed is circular in cross section from the beginning | 1. The cambial ring formed is wavy in the beginning and later becomes circular |
| 2. The cambial ring is partially primary (fasicular cambium) and partially secondary (Interfasicular cambium) in origin | 2. The cambial ring is completely secondary in origin |
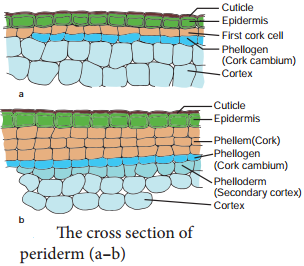
| 3. Generally, periderm originates from the cortical cells (extrastelar in origin) | 3. Generally, periderm originates from the pericycle.(intrastealar in origin) |
| 4. More amount of cork is produced as stem is aboveground | 4. Generally, less amount of cork is produced as root is underground |
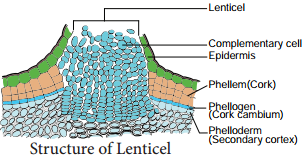
| 5. Lenticels of periderm are prominent | 5. Lenticels of periderm are not very prominent |
Most of the dicotyledonous roots show secondary growth in thickness, similar to that of dicotyledonous stems. Certain dicotyledonous roots do not show secondary growth. The secondary vascular tissues originate as a result of the cambial activity. The phellogen gives rise to the periderm.
Secondary growth takes place in all dicotyledonous woody plants. The root increases in girth by the activity of stelar and extrastelar cambium.
This is followed by periclinal division of the cells of pericycle present against protoxylem to form multiple layers of cells, which are joined by cambial cells derived from conjunctive tissues and together they make a complete cambium ring. Thus, the correct answer is option D.
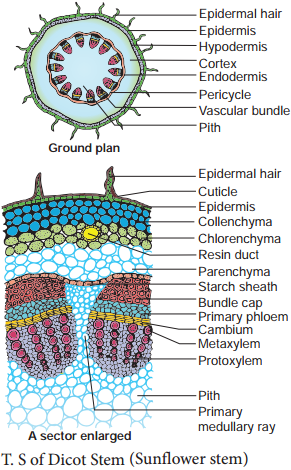
In a dicot stem, secondary growth occurs both in the stele and cortex. The process occurs simultaneously but is caused by separate strips of secondary meristem. In the stele, secondary growth is initiated by vascular cambium, while in the cortex, it is initiated by cork cambium.
In botany, secondary growth is the growth that results from cell division in the cambia or lateral meristems and that causes the stems and roots to thicken, while primary growth is growth that occurs as a result of cell division at the tips of stems and roots, causing them to elongate, and gives rise to primary tissue. There are two types of lateral tissues involved in secondary growth, namely, vascular cambium and cork cambium.
In general, monocots do not undergo secondary growth. If they do increase in girth (like palm trees and yucca plants), it does not result in the development of a secondary xylem and phloem, since monocots don’t have vascular cambium.
Difference Between the Secondary Growth in Dicot Stem and Dicot Root. The growth in thickness by the activity of secondary tissues is called secondary thickening. It involves stelar growth by the activity of vascular cambial ring and extra stelar growth by the activity of cork cambium.
Initiation of secondary growth takes place in the zone of maturation soon after the cells stop elongating there. The vascular cambium differentiates between the primary xylem and phloem in this zone and pericycle cells divide simultaneously with the procambium initials.
A process of formation of secondary tissues due to activity of vascular cambium and cork cambium for increasing thickness or girth or diameter of plant is termed as secondary growth.
Bougainvillea is a member of the Nyctaginaceae and is an example of a dicotyledonous stem which displays anomalous secondary growth. In this TS, near the centre of the stem, you will see some primary vascular bundles embedded in lignified pith parenchyma.
Secondary growth is characterized by an increase in thickness or girth of the plant, and is caused by cell division in the lateral meristem. Secondary vascular tissue is added as the plant grows, as well as a cork layer. The bark of a tree extends from the vascular cambium to the epidermis.
The increase in length of the shoot and the root is referred to as primary growth. It is the result of cell division in the shoot apical meristem. Secondary growth is characterized by an increase in thickness or girth of the plant.
The primary root grows vertically downwards into the soil. Smaller lateral roots known as the secondary roots are produced on the primary root. The secondary roots in turn produce tertiary roots. These roots grow in various directions and help in fixing the plant firmly into the soil.
Secondary growth is the growth in thickness due to the formation of secondary tissues by lateral meristems. Secondary growth does not occur in monocots because monocots do not possess vascular cambium in between the vascular bundles.
Secondary growth is the outward growth of the plant, making it thicker and wider. Secondary growth is important to woody plants because they grow much taller than other plants and need more support in their stems and roots. Lateral meristems are the dividing cells in secondary growth, and produce secondary tissues.
The process of secondary growth is controlled by the lateral meristems, and is similar in both stems and roots. Lateral meristems include the vascular cambium and, in woody plants, the cork cambium (cambium is another term for meristem).
Secondary xylem is a complex tissue that consists not only of non-living supporting and conducting cells but also of important living components (rays and axial wood parenchyma) which, with those in the secondary phloem, comprise a three-dimensional symplastic pathway through which photosynthate and other essential.
Lateral meristems are known as secondary meristems because they are responsible for secondary growth, or increase in stem girth and thickness. Meristems form anew from other cells in injured tissues and are responsible for wound healing.
Plant growth from lateral meristems such as the vascular cambium and cork cambium. This growth thickens plants and creates wood and bark (only in woody plants). Allows for taller, stronger plants, more branching and reproduction, and more conduction of fluids.
An example of a secondary meristem is the lateral meristem (e.g. cork cambium and accessory cambia). Being meristematic, the secondary meristem is comprised of undifferentiated (or partially differentiated), actively dividing cells.
The vascular cambium is responsible for increasing the diameter of stems and roots and for forming woody tissue. The cork cambium produces some of the bark. Cell division by the cambium produces cells that become secondary xylem and phloem.
In botany, secondary growth is the growth that results from cell division in the cambia or lateral meristems and that causes the stems and roots to thicken, while primary growth is growth that occurs as a result of cell division at the tips of stems and roots, causing them to elongate, and gives rise to primary tissue.
Secondary growth is the growth in thickness due to the formation of secondary tissues by lateral meristems. Secondary growth does not occur in monocots because monocots do not possess vascular cambium in between the vascular bundles.
In particular, secondary growth is substantial for constant plant growth and the remodeling of body structures.
As an important meristem involved, the vascular cambium forms a cylindrical domain below the organ surface producing tissues for long-distance transport and mechanical support: wood (xylem) and bast (phloem). Pericycle forms the boundary of stele and encloses vascular bundles and pith. It is a primary structure and is not formed as a result of secondary growth.